This article originally appeared at https://jpet.aspetjournals.org/content/377/2/273
Authors: Bárbara Betônico Berg, Jaqueline Silva Soares, Isabela Ribeiro Paiva, Barbara Maximino Rezende, Milene Alvarenga Rachid, Stêfany Bruno de Assis Cau, Thiago Roberto Lima Romero, Vanessa Pinho, Mauro Martins Teixeira and Marina Gomes Miranda e Castor
Journal of Pharmacology and Experimental Therapeutics May 2021, 377 (2) 273-283; DOI: https://doi.org/10.1124/jpet.120.000479
Abstract
Cannabidiol (CBD) is a highly lipidic phytocannabinoid with remarkable anti-inflammatory effects. The aim of this study was to evaluate CBD’s effects and mechanisms of action in the treatment of mice subjected to acute graft-versus-host disease (aGVHD). aGVHD was induced by the transplantation of bone marrow cells and splenocytes from C57BL-6j to Balb-c mice. The recipient mice were treated daily with CBD, and the treatment reduced mouse mortality by decreasing inflammation and injury and promoting immune regulation in the jejunum, ileum, and liver. Analysis of the jejunum and ileum showed that CBD treatment reduced the levels of C-C motif chemokine ligand (CCL) 2, CCL3, CCL5, tumor necrosis factor α, and interferon γ (IFNγ). CCL3 and IFNγ levels were also decreased in the liver. Mechanistically, CBD also increased the number of cannabinoid receptor type 2 (CB2) receptors on CD4+ and forkhead box P3+ cells in the intestine, which may explain the reduction in proinflammatory cytokines and chemokines. Antagonists of the CB2 receptor reduced the survival rates of CBD-treated mice, suggesting the participation of this receptor in the effects of CBD. Furthermore, treatment with CBD did not interfere with the graft-versus-leukemia response. CBD treatment appears to protect aGVHD mice by anti-inflammatory and immunomodulatory effects partially mediated by CB2 receptor interaction. Altogether, our study suggests that CBD represents an interesting approach in the treatment of aGVHD, with potential therapeutic applications in patients undergoing bone marrow transplantation.
SIGNIFICANCE STATEMENT This study provides for the first time a mechanism by which cannabidiol, a phytocannabinoid with no psychoactive effect, induces immunomodulation in the graft-versus-host disease. Enhancing intestinal cannabinoid receptor type 2 (CB2) receptor expression on CD4+ and forkhead box P3+ cells and increasing the number of these regulatory cells, cannabidiol decreases proinflammatory cytokines and increases graft-versus-host disease mice survival. This effect is dependent of CB2 receptor activation. Besides, cannabidiol did not interfere with graft-versus-leukemia response, a central response to avoid primary disease relapse.
Introduction
The main complication after allogeneic hematopoietic cell transplantation (alloHSCT) is graft-versus-host disease (GVHD), a systemic inflammatory condition secondary to alloHSCT that leads to significant morbidity and mortality in humans (Major-Monfried et al., 2018). AlloHSCT is the transplantation of cells between two genetically nonidentical individuals (Beatty et al., 1995; Ljungman et al., 2006) and is a curative therapy for several malignant and nonmalignant diseases, including leukemia (Gale, 1981), lymphoma (Curtis et al., 1997; Hartmann et al., 1997), thalassemia (Lucarelli et al., 1990), aplastic anemia (Locasciulli et al., 2007), and autoimmune diseases, such as lupus (Marmont, 1994; Gladstone et al., 2017).
GVHD occurs because of histoincompatibilities identified by T cells that react against host tissues causing extensive damage (Ho and Soiffer, 2001). Classically, acute GVHD (aGVHD) occurs within 100 days after transplantation, and chronic GVHD occurs after this period. However, nowadays, aGVHD is defined mainly by erythematous rash, gastrointestinal symptoms, cholestatic hepatitis, immune suppression, T cell infiltration, and tissue apoptosis (Zeng, 2018). The pathophysiology of aGVHD is complex and involves several inflammatory cytokines whose release starts during the conditioning regimen (irradiation and/or chemotherapy), leading to a proinflammatory environment that promotes T cell activation, proliferation, and differentiation against target organs (Schroeder and DiPersio, 2011). About 50% of patients with GVHD will die because of disease complications, and this is matched only by aGVHD’s high incidence rate among patients who undergo alloHSCT (Barton-Burke et al., 2008; Yang et al., 2017). Although alloHSCT leads to a higher chance of developing aGVHD, it also provides a stronger graft-versus-tumor (GVT) reactivity by which the new immune system recognizes and mediates the immunologic eradication of reminiscent cancer cells (Noriega et al., 2015; van Bergen et al., 2017). Therefore, GVT reactivity represents a strong plea in the recommendation of alloHSCT as a curative therapy for malignant conditions (Kolb, 2008).
The current treatment of aGVHD includes high doses of steroids, thus promoting host immune suppression but also leading to several adverse effects, such as reduced GVT reactivity, so new therapeutic approaches are urgently needed (Devetten and Vose, 2004; Shlomchik, 2007; Devergie and Janin, 2008; (Poetker and Reh, 2010); Peppa et al., 2011; Jamil and Mineishi, 2015). Meanwhile, cannabinoids emerge as strong pharmacological candidates for several pathologies, such as multiple sclerosis (Kozela et al., 2011), colitis (Borrelli et al., 2009), and inflammatory bowel disease (Schicho and Storr, 2012), but their psychoactive effects restrict their widespread therapeutic use. Cannabidiol (CBD), an abundant phytocannabinoid of Cannabis sativa with well known anti-inflammatory effects, is not psychoactive (Häußermann and Wagner, 2017; Adams et al., 2018) and is currently in a phase II study for the treatment of GVHD in humans with reports of benefits from CBD treatment, such as increased survival (Yeshurun et al., 2015). Furthermore, although there have been attempts to identify CBD’s mechanisms of action in murine models (Khuja et al., 2019), several gaps remain regarding the alterations triggered by CBD in the host organism and immune system. Therefore, the aim of this study was to investigate how CBD treatment can lead to an increase in mouse survival and by what mechanisms it can modulate the immune response regarding murine aGVHD.
Materials and Methods
Animals.
All the experiments were performed on 8-week-old male Balb-c mice, and the donor group consisted of 8-week-old male C57BL/6j mice from the Center of Bioterism, Federal University of Minas Gerais, Belo Horizonte, Brazil. The animals were housed under a 12-hour light/dark cycle with food and water available ad libitum. The animal procedures were approved by the Ethics Committee on Animal Experimentation of the Federal University of Minas Gerais (337/2014) and were carried out according to the Guide for the Care and Use of Laboratory Animals. The mice were euthanized through cervical displacement.
The sample size was defined based on the sample variance of three different GVHD-induction experiments previously performed by our group (Castor et al., 2010)(Castor et al., 2011). For the sample size, we used the following equation: in which n represents the sample size, zα/2 is the relative value from the z table, e is the absolute error, and σ is the standard deviation. To obtain a reliability of 95%, we needed a sample size of five mice per group for our experiments.
in which n represents the sample size, zα/2 is the relative value from the z table, e is the absolute error, and σ is the standard deviation. To obtain a reliability of 95%, we needed a sample size of five mice per group for our experiments.
Disease Induction.
For bone marrow ablation, the mice were irradiated with 7 Gy of γ radiation before transplantation and received 3 × 107 splenocytes and 1 × 107 bone marrow cells intravenously either from the C57BL/6j donor mice (and thus developed GVHD) or from the Balb-c donor (and thus did not develop GVHD); the mice receiving cells from the Balb-c donor were considered the control group. The cells were acquired according to Castor et al. (2010). The mice were randomly divided into the groups. After induction, the mice were monitored daily using a clinical standard scoring system consisting of seven points: hunching/posture, activity, fur texture, weight loss, skin integrity, diarrhea, and occult blood in the feces. The scoring was given according to the grade of clinical manifestation, with 0 = normal, 1 = minor alteration, 1.5 = moderate alteration, and 2 = severe alteration, as described previously (Castor et al., 2010). To avoid animal suffering, we standardized 10 points as the cutoff point.
Drugs.
The mice receiving C57BL/6j cells (allogeneic transplant) were treated either with CBD (THC-Pharm, Frankfurt, Germany and STI-Pharm, Brentwood, UK) in doses of 10, 30, and 60 mg/kg or its vehicle (5% Tween80 in sterile PBS-1×). The participation of cannabinoid receptors was assessed by a treatment with AM251 (Tocris) 1 mg/kg or AM630 (Tocris) 1 mg/kg (35), being CB1 and cannabinoid receptor 2 (CB2) antagonists, respectively. The vehicle of both these antagonists contained 2.5% DMSO in PBS-1× and CBD’s vehicle. All drugs were administered by a daily intraperitoneal injection starting on the day of disease induction and continuing for the duration of the protocols.
Histopathology.
On day 7, the intestine and liver were fixed and stained with H&E. The entire extension of the jejunum and ileum walls and the liver were analyzed blindly by a fellow pathologist under an optical microscope (BX51; Olympus, Tokyo, Japan). Numerical values were assigned to changes observed in each of the intestinal layers (the mucosa, lamina propria, muscular, and serosa layers). The epithelium was scored as follows: 0 = without alterations, 1 = reactive and aspecific alterations, 2 = erosion or loss of the architecture of the region of crypts or the surface of the epithelium, and 3 = nuclear changes and hyperplasia in the crypt or surface epithelium, with or without evidence of ulceration and loss of globet cells. The other layers were scored as follows: 0 = normal aspect; 1 = discrete inflammatory cell infiltration; 2 = moderate inflammatory cell infiltration, swelling, and congestion; 3 = severe inflammatory infiltration and villous enlargement with swelling and congestion in the lamina propria; and 4 = ischemic necrosis and intense inflammatory alterations in the muscular and serosa layers. Each animal received a total score generated by the summation of the three individual scores (maximum index 9). In the liver, there were degenerative alterations in the parenchyma, and a numerical value was assigned to the changes observed: 0 = normal; 1 = discrete cytoplasmatic vacuolization and focal eosinophilia; 2 = diffuse vacuolization, alterations in hepatocyte form, and moderate nucleus alterations; and 3 = hepatocyte necrosis and diffuse vacuolization, alterations in hepatocyte form, and accentuated nucleus alterations. Inflammatory infiltration of the liver was scored as follows: 0 = none or rare, 1 = discrete presence of infiltrates in the periportal area, 2 = discrete or moderate presence of infiltrates in the periportal and intralobular areas, and 3 = accentuated presence of infiltrates in the periportal and intralobular areas. Each animal received a total score generated by the summation of the two individual scores (maximum index 6).
Bacterial Translocation.
Intraperitoneal wash was performed on the 7th day. The mice were anesthetized with 80 µl of ketamine/xylazine (2:1), and the washing was performed with 1 ml of a sterile solution of PBS 1×. The wash was incubated overnight in a Müller-Hilton agar medium at 37°C, and the number of colony-forming units (CFUs) was counted. The mean values of the CFUs are shown.
ELISA.
On the 7th day, the mice were euthanized, and 100 mg of the intestine and liver were collected for the ELISA assay. The samples were then processed following the manufacturer’s instructions (R&D System) and were read in a wavelength of 490 nm in a spectrophotometer, and the absorbance value was determined for each sample. We evaluated CCL2, CCL3, CCL5, TNFα, and IFNγ.
Flow Cytometry.
Seven days after disease induction, the intestine, spleen, liver, and bone marrow were collected and processed as previously described (Resende et al., 2017). Cells were counted, and a total of 1 × 106 cells were stained with the following antibodies: CD3 [Pacific Blue (RRID-17-A2), BioLegend], CD4 [PE-Cy7 (RRID-RM45), BioLegend], CD8 [FITC (RRID-53-6.7), BioLegend], CD25 [allophycocyanin (RRID-PC61), BioLegend], and CD28 [PE-A (RRID-37.51), BioLegend]. For cell surface markers, we used FoxP3 [PE-A (RRID-MF14), BioLegend], H2Db [AlexaFluor 647 (RRID-KH95), BioLegend], and H2Dd [PE (RRID-34,212), BioLegend) for the bone marrow to evaluate chimerization. A total of 100,000 events were considered in the FACS analysis for each sample, and the cells were acquired with a BD FACSCanto II cytometer and analyzed using FlowJo V10 software (BD Immunocytometry Systems, San Jose, CA).
Immunofluorescence.
At day 7 after transplant, intestinal slices were processed to immunofluorescence assay. The antibodies to CD4, CD8, FoxP3, and CB2 described on supplemental material (Supplemental Fig. 1) were used to investigate the presence of CB2 on these cells and whether cannabidiol treatment could induce CB2 expression on them. The nuclei were marked with 4,6-diamidino-2-phenylindole (DAPI) (D1306-Carlsbad; Life Technology). Slides were mounted using 60% glycerol. Zen microscope was used to acquire images with suitable filters for selectively detecting the fluorescence of FITC (green), AlexaFluor 488 (green), AlexaFluor 594 (red), PE (red), or DAPI (blue) (Gonzaga et al., 2017). For each tissue section, 10 regions were randomly captured by an independent experiment-blinded user at 100× magnification, totaling 50 pictures per group (n = 5). Images were analyzed using ImageJ software v 1.53c 1.41 (National Institutes of Health, public domain).
Graft-versus-Tumor Response.
On the day of GVHD induction, after radiation, the mice received murine mastocytoma P815+ cells intravenously (H-2d; American Type Culture Collection, Rockville, MD–RRID TIB-64) transduced with an EF1aGFP vector (National Institute of Cancer, Rio de Janeiro, Brazil) in a concentration of 5 × 104 cells. The mice were treated with a vehicle or CBD for 7 days. The spleen and lymph nodes (mesenteric and inguinal) were separately processed in the FACS analysis by flow cytometry (FacsScaibur). The number of tumor cells was evaluated by the percentage of GFP+ fluorescence.
Statistical Analysis.
Statistical analysis was performed with GraphPad Prism 8.0.2 software. Normality was tested by the Shapiro-Wilk test, and outliers were identified by the Grubbs test (α = 0.05). The samples were tested by one-way ANOVA followed by Fisher’s LSD post hoc test to address differences between groups. In the survival curves, the differences were assessed by the log-rank (Mantel-Cox) test. Samples were considered different when P < 0.05.
Results
Engraftment and Mouse Survival.
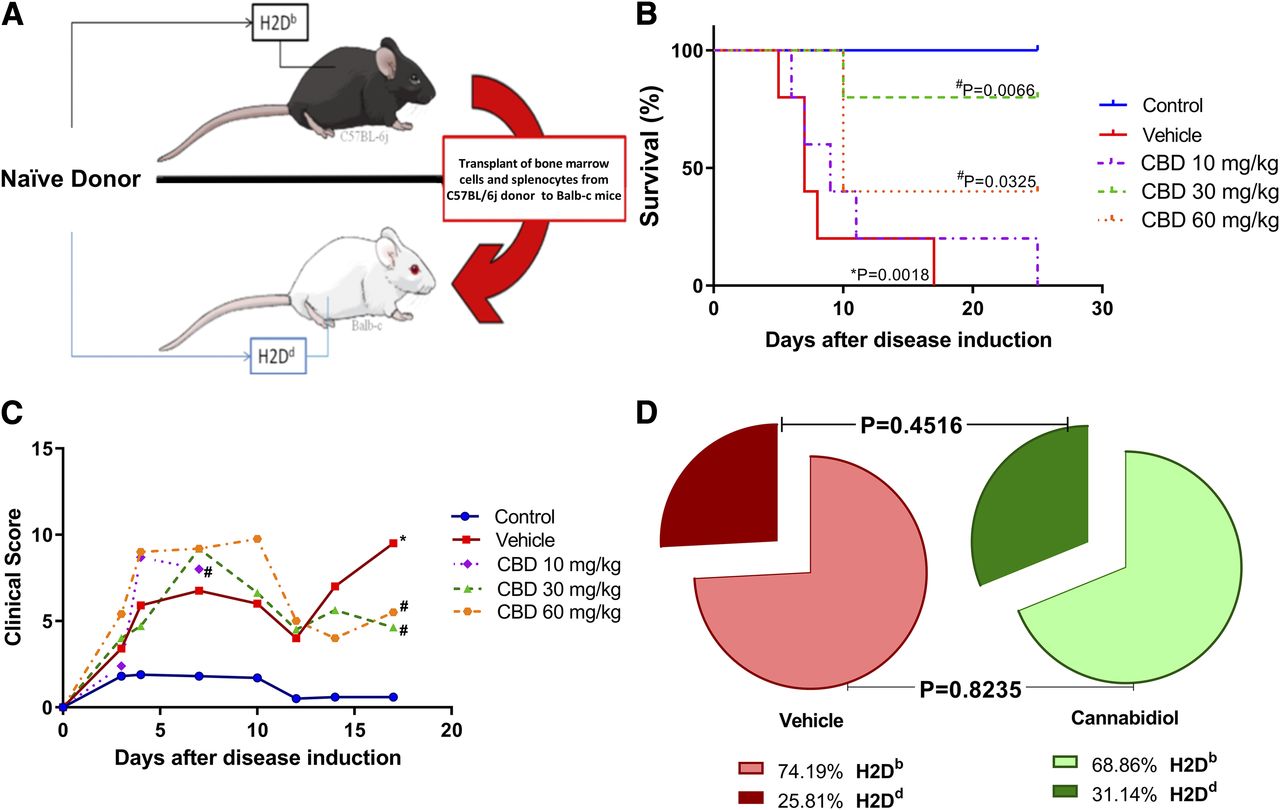
Major histocompatibility complex (MHC) disparities between animals led to the development of GVHD (Fig. 1A). The most effective dose of CBD was chosen by a daily treatment with 10, 30, or 60 mg/kg. Survival was assessed daily (Fig. 1B). We found no difference between CBD 10 mg/kg and the vehicle. Regarding the other doses, the CBD–30 mg/kg group displayed the highest survival (80%), with the CBD–60 mg/kg group showing a survival rate of 40%. CBD 30 mg/kg was clearly the most effective dose when compared with CBD 10 mg/kg (P = 0.0127), but there was no statistically significant difference between CBD 30 mg/kg and CBD 60 mg/kg (P = 0.2207). The clinical parameters of the mice were evaluated for 20 days and translated into a score (Fig. 1C). In this way, a dose of 30 mg/kg was chosen to be used for the next experimental procedures. During our experiments, the CBD–30 mg/kg group displayed a variation from 60% to 80% in survival, and the mortality in the vehicle group varied from 12 to 18 days.

{kind=link}
{kind=link}
Fig. 1.
Clinical aspects of experimental aGVHD. (A) Allogeneic transplant was performed from C57BL6j mice with H2Db phenotype to Balb-c mice with H2Dd phenotype. (B) Mice were treated with CBD 10 mg/kg, CBD 30 mg/kg, CBD 60 mg/kg, or vehicle, and their survival was assessed daily. (C) Daily assessment of mice through clinical score. (D) Bone marrow engraftment was verified in vehicle and CBD 30 mg/kg 7 days after the transplant through flow cytometry. * indicates P < 0.05 when compared with control group and # indicates P < 0.05 when compared with the vehicle. Data are expressed in dispersion with mean, and statistical difference was determined by Student’s t test for engraftment and log-rank (Mantel-Cox) test for survival. n = 5 for all groups
Furthermore, the bone marrow samples collected 7 days after disease induction revealed that the CBD–30 mg/kg treatment did not interfere with engraftment, as assessed by the percentages of the histocompatibility antigens H2Db and H2Dd in the CBD and vehicle groups. The vehicle group presented 75% of H2Db, and the mice treated with CBD presented 69% of H2Db (Fig. 1D).
Cellular Recruitment, Activation, and Damage.
The spleen, intestine, and liver were collected for analysis. Flow cytometry of the spleen (Supplemental Fig. 2) revealed no difference in the total number of lymphocytes (Fig. 2A) or in the number of CD8+ cells among the groups (Fig. 2C). Additionally, we found an increased number of CD4+ cells in the vehicle group when compared with the control group, but there was no difference between the CBD and the vehicle group in this respect (Fig. 2B). Moreover, we found no difference in the activation of CD4+ cells (Fig. 2D), but CD8+ cells were found to be increased in the vehicle group when compared with the control group. This profile was not altered by CBD treatment (Fig. 2E), and the number of CD4+, CD25+, and FoxP3+ cells, collectively known as Tregs, was not different between the groups (Fig. 2F).

{kind=link}
{kind=link}
Fig. 2.
Flow cytometry of mice spleen 7 days after disease induction. (A) Total lymphocytes; (B) CD3+CD4+ cells; (C) CD3+CD8+ cells; (D) CD3+CD4+CD28+; (E) CD3+CD8+CD28+; and (F) CD4+CD25+FoxP3+. * indicates P < 0.05 when compared with control group and # when P < 0.05 when compared with vehicle. Data are expressed in dispersion with mean, and statistical difference was determined by ANOVA with Fisher’s LSD post hoc. n = 5 for all groups
The H&E slices of the small intestine revealed that CBD treatment reduced damage to the tissue, preserving intestinal crypts, reducing inflammatory infiltration, preventing edema, and improving the general tissue score when compared with the vehicle group (Fig. 3, A–D). Supportive of this improvement in the small intestine, intraperitoneal washing also revealed that although mice from the vehicle group, as compared with the control group, had an increased number of CFUs translocating to the peritoneal cavity, CBD treatment prevented translocation, protecting the intestinal barriers (Fig. 3E).

{kind=link}
{kind=link}
Fig. 3.
Intestines stained with H&E, (A) control, (B) vehicle, and (C) CBD; stars represent edema, the dashed circle represents intestinal crypt, and arrows point to mononuclear infiltrate. (D) Quantification of the histopathological score; the preservation of the intestine morphology is translated in (E) reduced bacteria translocation. Data are expressed in dispersion with mean, and statistical difference was determined by ANOVA with Fisher’s LSD post hoc; n = 5 for all groups.
Flow cytometry of the small intestine (Supplemental Fig. 3) revealed an increased number of total lymphocytes in the CBD group in comparison with the vehicle group; however, the vehicle group had not an increased number of total lymphocytes in comparison with the control group (Fig. 4A).

{kind=link}
{kind=link}
Fig. 4.
Flow cytometry and ELISA of mice small intestines 7 days after disease induction. (A) Total lymphocytes; (B) CD3+CD4+ cells n = 5/5/5; (C) CD3+CD8+ cells n = 5/5/5; (D) CD3+CD4+CD28+ n = 5/5/4; (E) CD3+CD8+CD28+ n = 5/5/4; (F) CD4+CD25+FoxP3+ n = 5/5/5 cells; (G) CCL2, (H) CCL3, (I) CCL5, (J) TNFα, (K) IFNγ, and (L) IL-10. Data are represented in dispersion graphs with mean; n = 5 for all groups, except when specified. * indicates P < 0.05 when compared with control group and # when P < 0.05 when compared with vehicle. Data are expressed in dispersion with mean, and statistical difference was determined by ANOVA with Fisher’s LSD post hoc.
Both CD4+ cells (Fig. 4B) and CD8+ cells (Fig. 4C) were increased in the CBD group when compared with the vehicle group, but there was no difference between the vehicle and the control group. Interestingly, there was no difference in the activation of CD4+ cells between the CBD and the vehicle group, but in the latter group, the activation of CD4+ cells was increased in comparison with the control group (Fig. 4D). As for CD8+ cells, the vehicle group revealed an increase in activation when compared with the control group, and CBD did not prevent this activation (Fig. 4E). Interestingly, the CBD group displayed a higher number of Tregs when compared with the vehicle group, and the latter group showed no difference from the control group in this respect (Fig. 4F). Next, at the same time point, we verified the cytokine and chemokine profiles of the mice. The vehicle group displayed an increase in CCL2, CCL3, TNFα, IFNγ, and IL-10, whereas CBD, surprisingly, led to a reduction in CCL2, CCL3, CCL5, TNFα, IFNγ, and IL-10 in the intestine (Fig. 4, G–L).
The same parameters were evaluated in the mice liver 7 days after disease induction. Liver slices stained with H&E showed that CBD, as compared with the vehicle, was able to reduce inflammatory infiltration in the periportal and intralobular regions near the centrilobular veins of the liver (Fig. 5, A–D). In addition, ELISA in the liver showed that CBD decreased the levels of CCL2, CCL3, and IFNγ (Fig. 5 E, F, and I). Flow cytometry of the liver (Supplemental Fig. 4) revealed an increased number of lymphocytes in the CBD group when compared with the vehicle group (Fig. 5K), and this was also the case for CD4+ cells (Fig. 5L). However, we found no difference in CD8+ cells among the groups (Fig. 5M). Nevertheless, when activation was assessed by CD28+ staining, the CD4+ cells were found to be increased in the vehicle group when compared with the control group, whereas in the CBD group the activation was higher than in the vehicle group (Fig. 5N). The CD8+ cells presented the same low level of activation in all groups (Fig. 5O). Moreover, in the liver, the CBD group did not display a difference in the number of Treg cells when compared with the vehicle group, and neither did the vehicle group when compared with the control group (Fig. 5P).

{kind=link}
{kind=link}
Fig. 5.
Liver analysis. H&E from groups (A) control, (B) vehicle, and (C) CBD; dark arrows represent mononuclear infiltrate. (D) Quantification of histopathological score; ELISA of (E) CCL2, (F) CCL3, (G) CCL5, (H) TNFα, (I) IFNγ, and (J) IL-10; flow cytometry displaying (K) total lymphocytes; (L) CD3+CD4+ cells; (M) CD3+CD8+ cells; (N) CD3+CD4+CD28+ cells; (O) CD3+CD8+CD28+ cells; and (P) CD4+CD25+FoxP3+ cells. * indicates P < 0.05 when compared with control group and # when P < 0.05 when compared with vehicle. Data are expressed in dispersion with mean, and statistical difference was determined by ANOVA with Fisher’s LSD post hoc; n = 5 for all groups.
Graft-versus-Leukemia Response.
The mice received P815GFP+ mastocytoma cells on the day of disease induction. After 7 days, the response was assessed by flow cytometry. The control group received only tumor cells and a syngeneic transplant. In this group, we observed an increase in P815GFP+ cells in the spleen and lymph nodes, indicating tumor growth (Fig. 6, A and B). The vehicle group received tumor cells and allogeneic bone marrow and splenocytes, leading to a reduction in the number of GFP+ tumor cells compared with the control group (Fig. 6, A and B). In the same way, CBD treatment did not interfere with the allograft response against tumor cells (P815GFP+) since the presence of tumor cells was severely reduced in the spleen (Fig. 6A) and lymph nodes (Fig. 6B) of the mice that received CBD when compared with the mice in the control group, preserving the graft-versus-tumor response.

{kind=link}
{kind=link}
Fig. 6.
Flow cytometry of mice spleen and lymph nodes (LNs) 7 days after disease induction and P815GFP+ cells transplant. (A) Percentage of P815+ cells in lymph nodes. (B) Percentage of P815+ cells in spleen. Absolute number of P815+ cells are represented in dispersion graphs next to flow-cytometry shards; n = 5 for all groups.
CBD’s Effect Is Dependent on the CB2 Receptor.
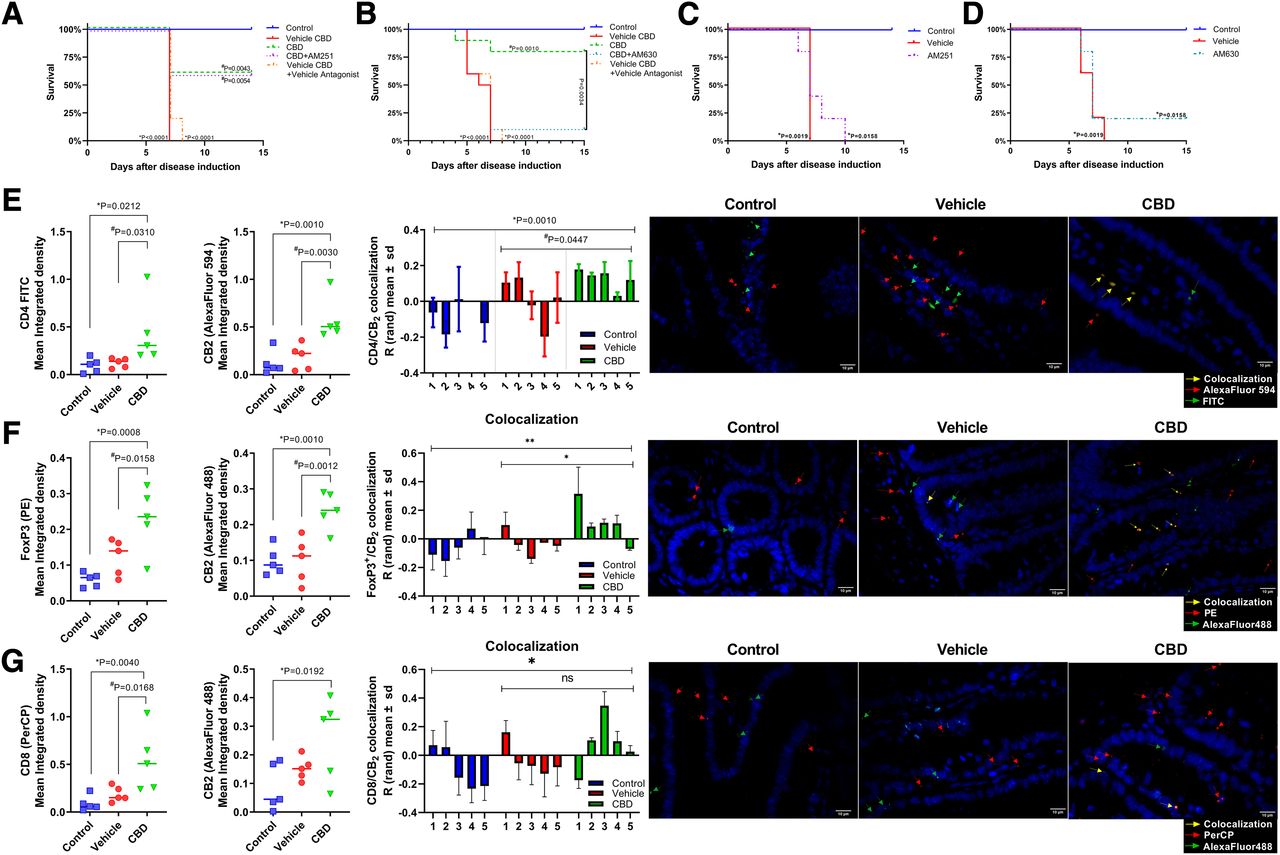
The mice were treated with CBD, CBD + AM630, CBD + AM251 or AM630, and AM251 alone, with AM630 being a selective antagonist of the CB2 receptor and AM251 being a selective antagonist of the CB1 receptor, and the survival of the mice was assessed. Interestingly, the mice that received AM251 with CBD survived, as did the mice from the CBD group (Fig. 7 A). Furthermore, both the vehicle group and the vehicle CBD + vehicle antagonist group were different from the control group (P < 0.001). The CBD group was different from its vehicle (P = 0.0043) but was not different from the CBD + AM251 group, and this last group was different from the vehicle CBD + vehicle antagonist group (P = 0.0054).

{kind=link}
{kind=link}
Fig. 7.
Role of CB2 in the protective effect of CBD. (A) CBD in the presence of CB1 antagonist; (B) CBD in the presence of CB2 antagonist; (C) treatment with CB1 antagonist without CBD; (D) treatment with CB2 antagonist without CBD. (E) CD4 and CB2 mean integrated density, colocalization of CD4/CB2, and representative figures of groups (CD4 FITC, CB2 AlexaFluor 594, and DAPI); (F) CD8 and CB2 mean integrated density, colocalization of CD8/CB2, and representative figures of groups (CD8 PerCP, CB2 AlexaFluor 488, and DAPI); (G) FoxP3 and CB2 mean integrated density, colocalization of FoxP3/CB2, and representative figures of groups (FoxP3, CB2 AlexaFluor 488, and DAPI). Differences between groups were assessed with log-rank (Mantel-Cox) test for P < 0.05; n = 10/10/5/5 for survival and ANOVA with Fisher’s LSD post hoc; n = 5 for all groups in immunofluorescence. ns, not significant.
Surprisingly, we found that the mice treated with AM630 together with CBD exhibited a lower survival rate than the mice treated with CBD alone (Fig. 7B). Moreover, the vehicle group’s survival was significantly different from the control group’s (P < 0.001); the same was found when the control group was compared with the vehicle CBD + vehicle AM630 group (P < 0.001). There was no difference between the vehicle CBD + vehicle AM630 group and the CBD + AM630 group. Finally, the CBD + AM630 group was different from the CBD group (P = 0.034). Additionally, the mice treated with either AM251 (Fig. 7C) or AM630 (Fig. 7D) were not different from their respective vehicle groups and did not display the same increase in survival as the mice that received CBD treatment.
To verify CB2 receptor expression in the target organs, we performed an immunofluorescence assay in the intestine, a major target organ of GVHD. CB2 expression is not altered by GVHD induction. However, CBD treatment increases CB2 receptor expression in the intestinal cells (Fig. 7E). As FACS analysis, immunofluorescence showed an increased number of CD4+ cells in the CBD group compared with GVHD and control group (Fig. 7E). Positive colocalization of CD4 and CB2 staining was increased in CBD group in comparison with both control and GVHD groups (Fig. 7E; Supplemental Fig. 5). This result was also found regarding FoxP3 with CB2 staining (Fig. 7F; Supplemental Fig. 6). However, although CD8 was also increased in CBD when compared with the other groups, CB2 receptor expression was found to be increased only in comparison with control and not GVHD group (P = 0.1232), and there was no difference between CBD and GVHD groups remarking colocalization (Fig. 7 G; Supplemental Fig. 7). Pearson’s coefficient was found to be mainly positive in CD4/CB2 analysis (control: 0.197 ± 0.127; GVHD: 0.161 ± 0.103; CBD: 0.333 ± 0.143; with no difference between groups), CD8/CB2 analysis (control: −0.0393 ± 0.169; GVHD: 0.275 ± 0.162; CBD: 0.253 ± 0.311; with no difference between groups), and regarding FoxP3 (control: 0.119 ± 0.075; GVHD: 0.179 ± 0.081; CBD: 0.233 ± 0.081; with statistical difference between control and CBD: P = 0.0418).
Discussion and Conclusions
First, we evaluated three different doses of CBD (10, 30, and 60 mg/kg) based on the published literature (De Filippis et al., 2011; Campos et al., 2015; Vilela et al., 2015; Ahmed and Katz, 2016) to determine the most appropriate dose for the treatment of aGVHD. The mice treated with CBD 30 mg/kg, however, were not free from the disease, as assessed by the clinical score, but displayed a reduced disease progression and mortality when compared with the vehicle group. Not only that, but when chimerism was assessed, CBD did not affect bone marrow engraftment, as the animals recovered to display the same percentage of bone marrow cells as the animals in the vehicle group, which was verified both in the bone marrow itself and in the spleen, a secondary lymphoid organ.
The absence of a functional immune system would have left the mice susceptible to opportunistic diseases and is proven to be of the highest importance when considering possible treatments for GVHD in humans (Jamil and Mineishi, 2015; Admiraal et al., 2017). Therefore, in the present study, the presence of active and proliferating T cells in the spleen reinforces that CBD does not lead to an immunosuppressive status, as it does not lead to a reduction in T CD4+ and CD8+ cells when compared with the vehicle group. Jamil and Mineishi (2015) highlighted the importance of not entirely suppressing GVHD because it is related to a higher GVT response, which will protect the recipient from reminiscent tumor cells. The presence of transplanted cells in secondary lymphoid organs is important for the protective effect of the GVT response; however, it also relates to aGVHD development (Tkachev et al., 2018). In our study, CBD treatment was able to protect target organs against damage without interfering with the GVT response. These first two results are of the utmost importance when considering CBD usage in both prevention and treatment of patients who undergo alloHSCT.
CD4+ cells are responsible for generating both thymic-derived and inducible Tregs (Ferrara et al., 2017). Tregs are characterized by the presence of CD4+, CD25+, and FoxP3+ markers and promote tolerability, suppressing alloreactive lymphocytes. This regulation is key to aGVHD suppression/reduction (Blazar et al., 2012). Like our findings, the results reported by Edinger et al. (2003) of an experimental model of Treg transfer showed that even though Tregs are able to reduce aGVHD, they did not affect the GVT response, probably because of cytolytic effector T cells, especially CD8+ cells in the lymphoid organs (Schneidawind et al., 2013). CD8+ cells or T cytotoxic cells are vital to antitumoral immunologic responses (Uri et al., 2018). In this regard, we attested that CBD treatment does not reduce the migration of CD4+ or CD8+ cells to lymphoid and target organs. In our study, the main organ evaluated was the intestine, which is a major target organ of aGVHD and is regarded as one of the main environments for antigen presentation, acting as a barrier against commensal microbiota (Peled et al., 2016). CBD protected the intestinal barriers by preventing the migration of bacteria into the peritoneal cavity, whereas in the vehicle group, the intestinal barrier was inefficient in preventing this translocation; owing to intestinal damage, bacteria were found in the peritoneal cavity (Fig. 3E).
In the intestine, CBD led to an increase in CD4+ and CD8+ populations, although there was no increase in the activation status of these cells. Another important difference between CBD and the vehicle is that CBD promotes Treg cells in the intestine, a factor that may elucidate the reduction in intestinal damage and the level of proinflammatory cytokines (TNFα and IFNγ) and chemokines (CCL2, CCL3, and CCL5). The chemokines CCL2, CCL3, and CCL5 are mandatory for recruitment of inflammatory cells to the target organs (Castor et al., 2010; Blazar et al., 2012; Ferrara et al., 2017). In this way, we have investigated whether the recruitment process was impaired in CBD-treated mice. Using an intravital assay, the rolling and adhesion of inflammatory cells on intestinal microvasculature were examined. There was no difference between vehicle- and CBD-treated mice (Supplemental Fig. 8).
Based on the findings by Panoskaltsis-Mortari et al. (2004), effector cells migrate first to the intestine and later to the liver. We hypothesize that CD4+ activation and the further differentiation of CD4+ cells into FoxP3+ cells would also occur later in the liver and that, therefore, the number of CD4+ cells were not increased on the 7th day of the disease in vehicle. The timing of the disease in the liver explains why we found that CCL2, CCL3, and IFNγ were increased only in the vehicle group; interestingly, this increase was prevented by CBD. Tregs have been described to suppress cytokine production of both CD4+ and CD8+ effector cells (Helling et al., 2015), a process that elucidates our findings regarding the ELISA assay in the intestine. Additionally, the presence of Tregs is necessary for the promotion of immune tolerance, preventing an overwhelming graft immune response after alloHSCT (Becker et al., 2007; Beres and Drobyski, 2013). To our knowledge, this is the first time that CBD has been shown to increase CD4+, CD25+, and FoxP3+ cells in an experimental model of aGVHD, an effect that elucidates part of the modulatory mechanism of the inflammatory response and the increase in mouse survival and organ protection.
Another interesting mechanism of CBD is that in our model the CB2 but not the CB1 receptor is partially responsible for mouse survival, and we believe that it may also be responsible for the immunomodulatory effects of CBD. Both receptors are classified as cannabinoid receptors, as they are sensitive to both endogenous cannabinoids and some phytocannabinoids; however, they unleash different effects in the host organism. The CB1 receptor is present mainly in the central nervous system, whereas the CB2 receptor is more abundant in the peripheral nervous system and immune cells (Adams et al., 2018). The presence of CB2 in immune cells is correlated with its modulatory effects, including cytokine inhibition and reduction in antigenic presentation (Ehrhart et al., 2005)(Horváth et al., 2012). CB2 receptor is expressed on nearly all immune cells, and its absence on donor CD4 + or CD8 + T cells or administration of a selective CB2 pharmacological antagonist exacerbated acute GVHD lethality (Yuan et al., 2021). CBD has about 74% higher affinity for CB2 than for CB1 (Rosenthaler et al., 2014). In this way, we investigated the effect of CBD on CB2 receptor expression on intestine. CBD treatment was able to induce an increase of CB2 receptor expression on CD4+ and FoxP3+ cells in the intestine of mice subject to GVHD. The correlation between CD4 and CB2 but also that of FoxP3 and CB2 further reinforce our hypothesis that cannabidiol might stimulate FoxP3 differentiation by interaction with CB2 receptor. These findings reinforce CB2’s role in regulation of GVHD and suggest an interesting pharmacological therapeutic targeting.
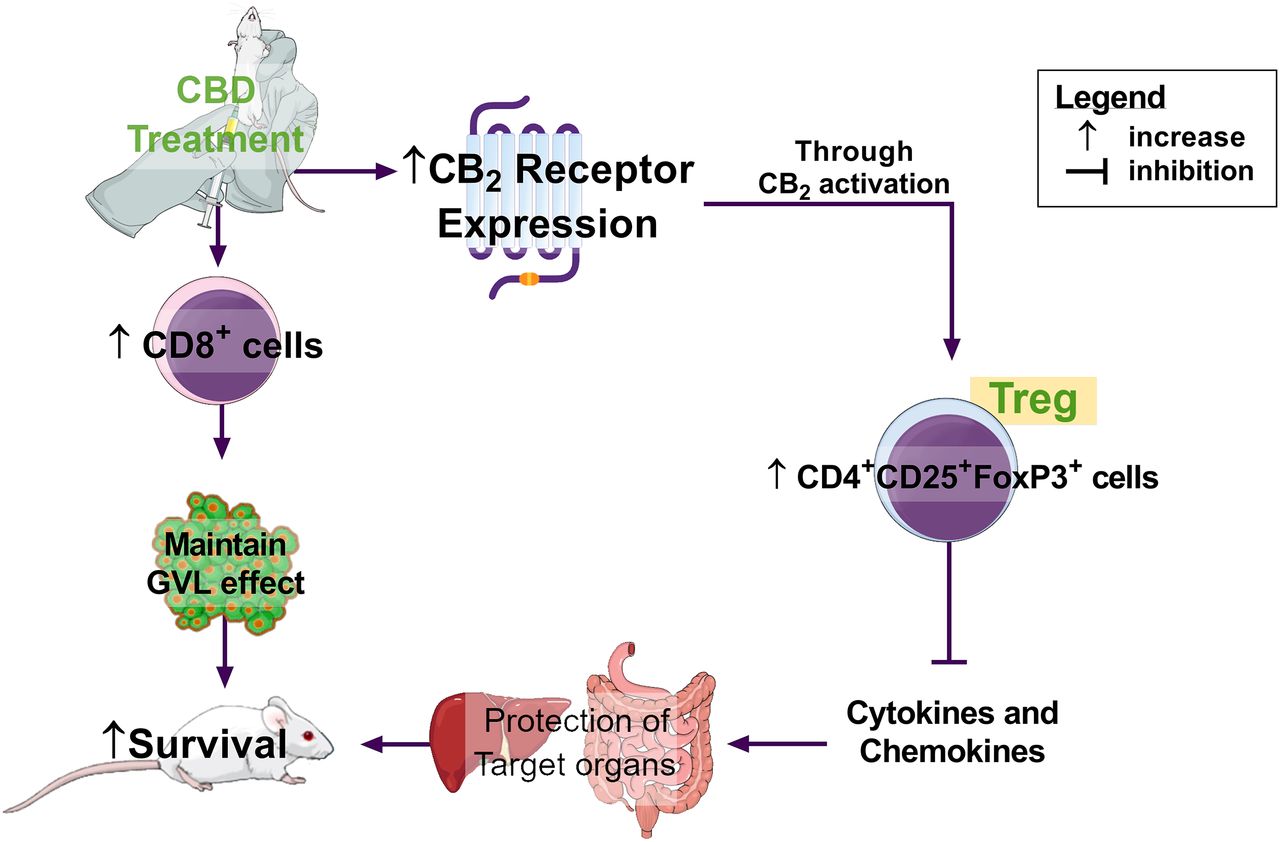
In the present study, the effects of CBD may be associated with CB2 activation. However, it is important to highlight that CBD can exert different effects upon a vast number of different receptors (Pertwee, 2006) and that, therefore, additional mechanisms may underlie the full modulatory effects of CBD. Lastly, in our model of aGVHD, CBD treatment was able to increase survival and reduce the damage of target organs. The increase of CB2 receptor in the intestine leads to an immunoregulatory environment with an enhanced number of Treg cells and a reduction in the major GVHD proinflammatory mediators. Moreover, CBD increases the number of CD8+ cells maintaining the graft-versus-leukemia effect. Therefore, it may be concluded that CBD represents an interesting approach in the treatment of aGVHD (Fig. 8).

{kind=link}
{kind=link}
Fig. 8.
Schematic representation of our findings. CBD treatment, partially by CB2 receptor activation, increased the number of Tregs in the intestines and reduced cytokine/chemokine release. Together these factors led to the protection of target organs and the increase in mice survival.
Acknowledgments
We would like to thank Aline C. Campos of the Escola de Medicina de Ribeirão Preto, Departamento de Farmacologia-USP for providing the cannabidiol for our experiments.
Authorship Contributions
Participated in research design: Berg, Romero, Pinho, Teixeira, Castor.
Conducted experiments: Berg, Soares, Paiva, Castor.
Contributed new reagents or analytic tools: Rezende, Cau, Romero, Pinho, Teixeira.
Performed data analysis: Berg, Soares, Rezende, Rachid, Castor.
Wrote or contributed to the writing of the manuscript: Berg, Pinho, Teixeira, Castor.
Footnotes
- Received December 21, 2020.
- Accepted February 22, 2021.
- The work was supported by grants from Coordenação de Aperfeiçoamento de Pessoal do Ensino Superior (CAPES): Conselho Nacional de Desenvolvimento Ciêntífico e Tecnológico (CNPQ) [Grant 444992-2014/6], Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) [Grant APQ-01307-14], and Pró-Reitoria de Pesquisa da UFMG (PRPq) [Grant 307-05/2016].
- The authors declare no conflict of interest in this study.
- This work was previously presented in the following scientific meetings: 4th International Symposium on Inflammatory Diseases (INFLAMMA IV), International Congress In Cannabinoid Medicine (CannX), XXVI Encontro de Pesquisa em Fisiologia and Farmacologia; Brazilian Congress of Pharmacology and Experimental Therapeutics (SBFTE), and The 5th Congress on Controversies in Stem Cell Transplantation and Cellular Therapies (COSTEM), with the work being rewarded in the last two.
- https://doi.org/10.1124/jpet.120.000479.
- ↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.
Abbreviations
aGVHDacute GVHDalloHSCTallogeneic hematopoietic cell transplantationCB1cannabinoid receptor type 1CB2cannabinoid receptor type 2CBDcannabidiolCCLC-C motif chemokine ligandCFUcolony-forming unitDAPI4,6-diamidino-2-phenylindoleFACSfluorescence-activated cell sortingFITCfluorescein isothiocyanateFoxforkhead boxGVHDgraft-versus-host diseaseGVTgraft-versus-tumorIFNγinterferon γILinterleukinLSDleast significant differencePEPhycoerythrinPerCPPeridinin-chlorophyll-proteinTNFαtumor necrosis factor αTregregulatory T cell
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics
References
- ↵
- Adams J,
- Unett D,
- Anthony T,
- Gatlin J, and
- Gaidarov I
- ↵
- Admiraal R,
- de Koning CC,
- Lindemans CA,
- Bierings MB,
- Wensing AM,
- Versluys AB,
- Wolfs TFW,
- Nierkens S, and
- Boelens JJ
- ↵
- Ahmed W and
- Katz S
- ↵
- Barton-Burke M,
- Dwinell DM,
- Kafkas L,
- Lavalley C,
- Sands H,
- Proctor C, and
- Johnson E
- ↵
- Beatty PG,
- Mori M, and
- Milford E
- ↵
- Becker C,
- Kubach J,
- Wijdenes J,
- Knop J, and
- Jonuleit H
- ↵
- Beres AJ and
- Drobyski WR
- ↵
- Blazar BR,
- Murphy WJ, and
- Abedi M
- ↵
- Borrelli F,
- Aviello G,
- Romano B,
- Orlando P,
- Capasso R,
- Maiello F,
- Guadagno F,
- Petrosino S,
- Capasso F,
- Di Marzo V, et al.
- ↵
- Campos AC,
- Brant F,
- Miranda AS,
- Machado FS, and
- Teixeira AL
- ↵
- Castor Marina GM,
- Rezende Barbara M,
- Bernardes Priscila T. T.,
- Vieira Angélica T.,
- Vieira Érica L. M.,
- Arantes Rosa M. E.,
- Souza Danielle G.,
- Silva Tarcília A.,
- Teixeira Mauro M., and
- Pinho Vanessa
- ↵
- Castor MG,
- Rezende B,
- Resende CB,
- Alessandri AL,
- Fagundes CT,
- Sousa LP,
- Arantes RM,
- Souza DG,
- Silva TA,
- Proudfoot AE, et al.
- ↵
- Curtis RE,
- Rowlings PA,
- Deeg HJ,
- Shriner DA,
- Socíe G,
- Travis LB,
- Horowitz MM,
- Witherspoon RP,
- Hoover RN,
- Sobocinski KA, et al.
- ↵
- De Filippis D,
- Esposito G,
- Cirillo C,
- Cipriano M,
- De Winter BY,
- Scuderi C,
- Sarnelli G,
- Cuomo R,
- Steardo L,
- De Man JG, et al.
- ↵
- Apperley J.,
- Carreras E.,
- Gluckman E.,
- Gratwohl A., and
- Masszi T.
- Devergie A and
- Janin A
- ↵
- Devetten MP and
- Vose JM
- ↵
- Edinger M,
- Hoffmann P,
- Ermann J,
- Drago K,
- Fathman CG,
- Strober S, and
- Negrin RS
- ↵
- Ehrhart J,
- Obregon D,
- Mori T,
- Hou H,
- Sun N,
- Bai Y,
- Klein T,
- Fernandez F,
- Tan J, and
- Shytle RD
- ↵
- Ferrara JL,
- Smith CM,
- Sheets J,
- Reddy P, and
- Serody JS
- ↵
- Gale RP
- ↵
- Gladstone DE,
- Petri M,
- Bolaños-Meade J,
- Dezern AE,
- Jones RJ,
- Fine D, and
- Brodsky RA
- ↵
- Gonzaga Amanda C.R.,
- Campolina-Silva Gabriel H.,
- Hipácia Werneck-Gomes,
- Júnia D. Moura-Cordeiro,
- Letícia C. Santos,
- Germán A.B. Mahecha,
- Mônica Morais-Santos, and
- Cleida A. Oliveira
- ↵
- Hartmann O,
- Le Corroller AG,
- Blaise D,
- Michon J,
- Philip I,
- Norol F,
- Janvier M,
- Pico JL,
- Baranzelli MC,
- Rubie H, et al.
- ↵
- Häußermann K and
- Wagner U
- ↵
- Helling B,
- König M,
- Dälken B,
- Engling A,
- Krömer W,
- Heim K,
- Wallmeier H,
- Haas J,
- Wildemann B,
- Fritz B, et al.
- ↵
- Ho VT and
- Soiffer RJ
- ↵
- Horváth B,
- Magid L,
- Mukhopadhyay P,
- Bátkai S,
- Rajesh M,
- Park O,
- Tanchian G,
- Gao RY,
- Goodfellow CE,
- Glass M, et al.
- ↵
- Jamil MO and
- Mineishi S
- ↵
- Khuja I,
- Yekhtin Z,
- Or R, and
- Almogi-Hazan O
- ↵
- Kolb H-J
- ↵
- Kozela E,
- Lev N,
- Kaushansky N,
- Eilam R,
- Rimmerman N,
- Levy R,
- Ben-Nun A,
- Juknat A, and
- Vogel Z
- ↵
- Ljungman P,
- Urbano-Ispizua A,
- Cavazzana-Calvo M,
- Demirer T,
- Dini G,
- Einsele H,
- Gratwohl A,
- Madrigal A,
- Niederwieser D,
- Passweg J, et al., and European Group for Blood and Marrow
- ↵
- Locasciulli A,
- Oneto R,
- Bacigalupo A,
- Socié G,
- Korthof E,
- Bekassy A,
- Schrezenmeier H,
- Passweg J,
- Führer M, and Severe Aplastic Anemia Working Party of the European Blood and Marrow Transplant Group
- ↵
- Lucarelli G,
- Galimberti M,
- Polchi P,
- Angelucci E,
- Baronciani D,
- Giardini C,
- Politi P,
- Durazzi SM,
- Muretto P, and
- Albertini F
- ↵
- Major-Monfried H,
- Renteria AS,
- Pawarode A,
- Reddy P,
- Ayuk F,
- Holler E,
- Efebera YA,
- Hogan WJ,
- Wölfl M,
- Qayed M, et al.
- ↵
- Marmont AM
- ↵
- Noriega V,
- Martínez-Laperche C,
- Buces E,
- Pion M,
- Sánchez-Hernández N,
- Martín-Antonio B,
- Guillem V,
- Bosch-Vizcaya A,
- Bento L,
- González-Rivera M, et al., and Spanish Hematopoietic Stem Cell Transplantation and Cell Therapy Group (GETH)
- ↵
- Panoskaltsis-Mortari A,
- Price A,
- Hermanson JR,
- Taras E,
- Lees C,
- Serody JS, and
- Blazar BR
- ↵
- Peled JU,
- Hanash AM, and
- Jenq RR
- ↵
- Peppa M,
- Krania M, and
- Raptis SA
- ↵
- Pertwee RG
- ↵
- Poetker DM and
- Reh DD
- ↵
- Resende CB,
- Rezende BM,
- Bernardes PT,
- Teixeira GM,
- Teixeira MM,
- Pinho V, and
- Bittencourt H
- ↵
- Rosenthaler S,
- Pöhn B,
- Kolmanz C,
- Huu CN,
- Krewenka C,
- Huber A,
- Kranner B,
- Rausch WD, and
- Moldzio R
- ↵
- Schicho R and
- Storr M
- ↵
- Schneidawind D,
- Pierini A, and
- Negrin RS
- ↵
- Schroeder MA and
- DiPersio JF
- ↵
- Shlomchik WD
- ↵
- Tkachev V,
- Furlan SN,
- Potter EL,
- Zheng BH,
- Hunt DJ,
- Colonna L,
- Taraseviciute A,
- Carlson J,
- Betz K,
- Yu A, et al.
- ↵
- Uri A,
- Lühder F,
- Kerkau T, and
- Beyersdorf N
- ↵
- van Bergen CA,
- van Luxemburg-Heijs SA,
- de Wreede LC,
- Eefting M,
- von dem Borne PA,
- van Balen P,
- Heemskerk MH,
- Mulder A,
- Claas FH,
- Navarrete MA, et al.
- ↵
- Vilela LR,
- Gomides LF,
- David BA,
- Antunes MM,
- Diniz AB,
- Moreira FdeA, and
- Menezes GB
- ↵
- Yang F,
- Lu D,
- Hu Y,
- Huang X,
- Huang H,
- Chen J,
- Wu D,
- Wang J,
- Wang C,
- Han M, et al.
- ↵
- Yeshurun M,
- Shpilberg O,
- Herscovici C,
- Shargian L,
- Dreyer J,
- Peck A,
- Israeli M,
- Levy-Assaraf M,
- Gruenewald T,
- Mechoulam R, et al.
- ↵
- Yuan Cheng Yin,
- Zhou Vivian,
- Sauber Garrett,
- Stollenwerk Todd,
- Komorowski Richard,
- López Alicia,
- Tolón Rosa María,
- Romero Julian,
- Hillard Cecilia J., and
- Drobyski William R.